فصل سوم: ویژگیهای ساختمانی و عملی یاخته های عصبی
یاخته عصبی یا نورون واحد ساختمانی و عملی دستگاه عصبی است. نورون مانند هر سلول دیگر بدن دارای هسته سیتوپلاسم و غشاست آنچه در نظر اول به یک نورون جلب توجه میکند وجود تعداد زیادی انشعابات سیتوپلاسمی است که به صورت رشته هایی از جسم سلولی خارج شده اند؛ به طوری که حجم این رشته ها از حجم جسم سلولی بسیار زیادتر است. هر یک از این رشته ها یک تار عصبی نامیده می شود و اجتماع تعدادی تار عصبی که به وسیله غلافی پیوندی پوشیده شده اند یک عصب را می سازند تارهای عصبی متصل به جسم سلولی نورون شامل یک آکسون و تعدادی دندریت است. آکسون یک رشته منفرد است که در انتهای خود به شاخه هایی تقسیم میشود. دندریتها انشعابات زیادی دارند و معمولاً طول آنها کوتاه تر از آکسونهاست؛ ولی دندریتهای برخی از نورونها مانند یاخته های حسی که پیامهای پوست را به مراکز عصبی میآورند طویل و شبیه آکسونهاست.
شکل و ساختمان جسم سلولی نورون
بخش هسته دار یاخته عصبی به نامهای پریکاریون و سوما خوانده می شود. شکل و اندازه پریکاریون در نورونهای مختلف متفاوت است. بسیاری از آنها به شکلهای مدور، بیضوی ،هر می ذوزنقه ای و ستاره ای اند و قطر آنها از چند میکرون تا چند صد میکرون، متفاوت است. سیتوپلاسم سلول عصبی را نوروپلاسم می خوانند. در نوروپلاسم اندامکهای ریزی که در سایر سلولهای زنده وجود دارد یافت می شود. شبکه درون سیتوپلاسم با دانه های ریبوزوم که بر روی آن قرار دارند، میتوکندریهای فراوان دانه های لیزوزوم دستگاه گلژی، گویچه های چربی و مواد رنگی از جمله این اندامکها هستند که هر کدام اعمالی را به عهده دارند و بسیاری از آنها درون تارهای عصبی نیز یافت میشوند دانه های ریبوزوم و شبکه آندوپلاسمی در پروتئین سازی و نقل و انتقال آنها نقش اساسی دارند و در برخی نواحی دانه های درشتی را می سازند که رنگهای قلیایی را به خود جذب می کنند و اجسام نیسل خوانده میشوند. میتوکندریها انرژی لازم را برای زندگی یاخته فراهم می کنند. دانه های لیزوزوم محتوی آنزیمهایی هستند که میتوانند ذرات خارجی و مواد زاید را تجزیه و تخریب کنند. دستگاه گلژی در غشاسازی و بسته بندی ترشحات نورون شرکت دارد میکروسکوپ الکترونی وجود رشته های بسیار نازک (نوروفیبریل) و لوله های بسیار باریک نورو توبول را در نوروپلاسم نشان می دهد. این رشته ها و لوله ها نوعی اسکلت سیتوپلاسمی تشکیل میدهند که به حفظ شکل نورون و تارهای عصبی کمک می کند. هسته نورون تفاوت خاصی با هسته سایر سلولها ندارد. در درون آن شیره هسته (نوکلئوپلاسم ، یک یا چند هستک (نوکلئول) و توده های کروماتین دیده می شود.
نورونها از نظر طرز خارج شدن تارهای عصبی از جسم سلولی به سه گروه یک قطبی ، دو قطبی و چند قطبی تقسیم میشوند در نورونهای یک قطبی آکسون و دندریتها از یک ناحیه جسم سلولی خارج میشوند. نمونه این نورونها در عقده های نخاعی انسان دیده میشود که گیرنده های حسهای پیکری هستند. اغلب نورونهای بی مهرگان نیز یک قطبی اند در نورونهای دو قطبی آکسون از یک ناحیه و دندریتها از ناحیه دیگر جسم سلولی بیرون می آیند گیرنده های بویایی که در مخاط زرد بینی قرار دارند و برخی نورونهای شبکیه چشم از این نوع هستند. در نورونهای چند قطبی که تعداد آنها بیش از دو نوع دیگر است آکسون از یک ناحیه و دندریتها از چند ناحیه دیگر جسم سلولی خارج میشوند. در این نورونها تشخیص آکسون از دندریتها به سادگی میسر است؛ در حالی که در نورونهای یک قطبی و دو قطبی تشخیص آکسون از دندریتها آسان نیست نورونهای حرکتی شاخ پیشین نخاع و نورونهای پورکینیه (پورکنژ) مخچه و نورونهای هرمی شکل قشر مخ همگی از نوع چند قطبی ولی با شکلهای متفاوت اند. در بعضی نواحی دستگاه عصبی مرکزی نورونهای فاقد آکسون شناسایی شده است. این نورونها تحریک عصبی را فقط به نورونهای مجاور خود منتقل می کنند.
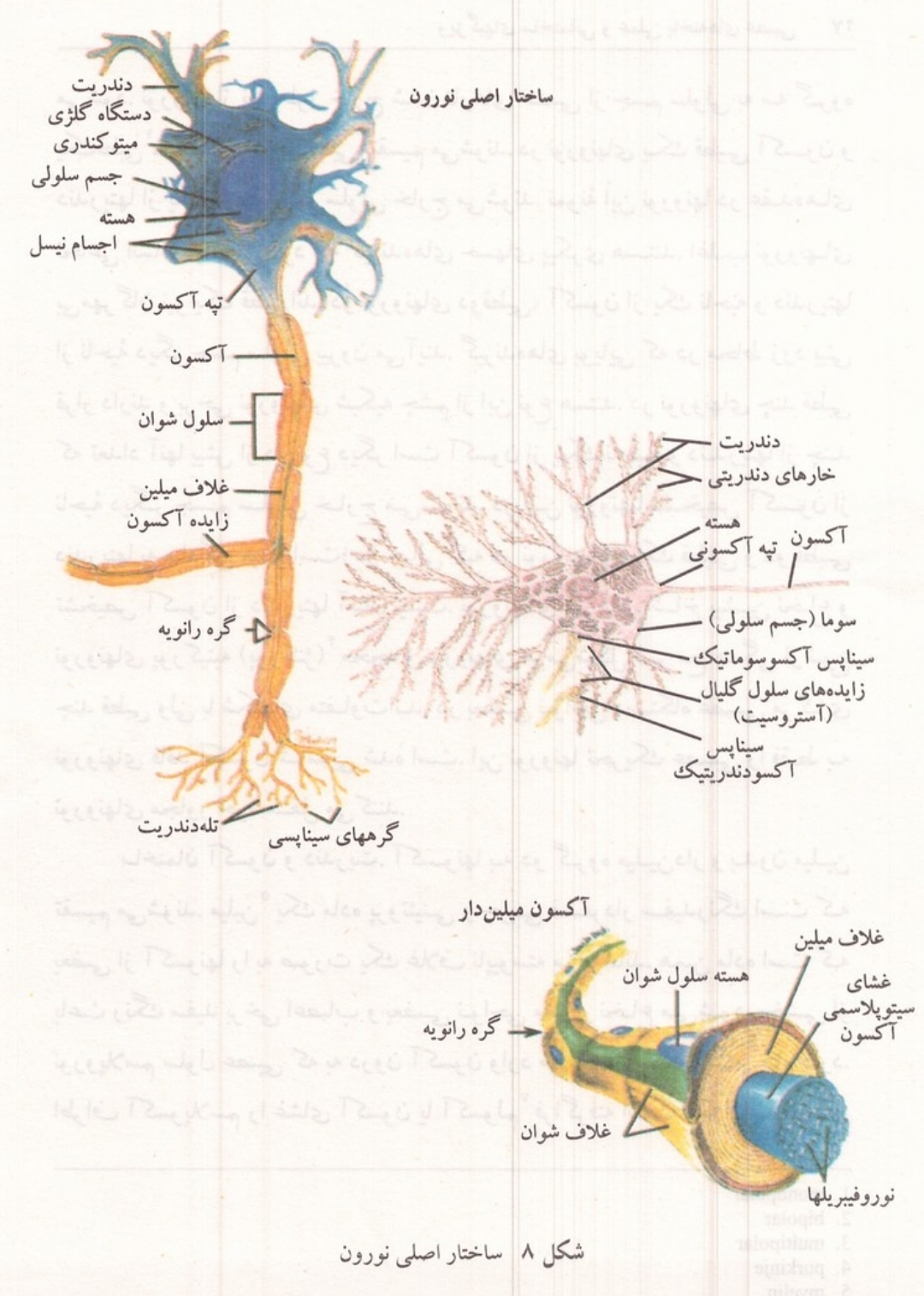
شکل ۸: ساختار اصلی نورون (چپ) و جزئیات آکسون میلین دار (راست)
ساختمان آکسون و دندریت
آکسونها به دو گروه میلین دار و بدون میلین تقسیم میشوند میلین یک ماده پروتئینی و چربی فسفردار سفیدرنگ است که بعضی از آکسونها را به صورت یک غلاف ناپیوسته میپوشاند. همین ماده است که باعث رنگ سفید برخی اعصاب و بعضی نواحی مغز و نخاع می شود. بخشی از نوروپلاسم سلول عصبی که به درون آکسون وارد میشود آکسوپلاسم نام دارد. اطراف آکسوپلاسم را غشای آکسون یا آکسولم فرا گرفته است که ادامه غشای نورون است. در اطراف آکسونهای بدون میلین ممکن است غلافهای پیوندی دیده شود. رنگ این تارها خاکستری و شبیه رنگ پریکاریون است. در آکسونهای میلین دار که قطر آنها بیش از تارهای بدون میلین است به فواصل مساوی گسستگیهایی در میلین دیده میشود. در این نواحی که گرههای را نویه خوانده می شوند غشای آکسون فاقد پوشش میلین است. فاصله بین دو گره را نویه مجاور در تارهای عصبی بر حسب قطر آنها بین ۰/۵ تا ۱/۵ میلی متر است. ابتدای آکسون که در مجاورت جسم سلولی قرار دارد مخروط یا پیاز آکسون نام دارد و فاقد میلین است. انتهای آکسون نیز قبل از ناحیه انشعاب آن میلین ندارد. میلین به صورت مارپیچی در اطراف آکسون ساخته میشود میلین در آکسونهای اعصاب پیرامونی به وسیله سلولهایی به نام شوان و در مراکز عصبی به وسیله یک نوع از بافت همبند عصبی به نام اولیگودندرو گلیا ساخته میشود مجموعه سلولهای شوان یا اولیگودندرو گلیا که میلین را ساخته اند به صورت غلافی تار عصبی میلین دار را فرامی گیرند. آکسونهای میلین دار فقط در اعصاب مهره داران وجود دارند و اعصاب بی مهرگان همگی از رشته های بدون میلین ساخته شده است.
دندریتها از واژه دندرون به معنی درخت شاخه های باریک زیادی دارند. قطر قاعده دندریت بیشتر از انتهای آن است و نازک ترین شاخه های دندریتها را فقط با میکروسکوپ الکترونی میتوان مشاهده کرد در درون دندریتها، میتوکندری شبکه درون سیتوپلاسم ریبوزومها و سایر ضمایم سیتوپلاسمی نورونها دیده می شود. سطح خارجی دندریتها دارای گیرنده های غشایی است که اطلاعات را از نورونهای دیگر دریافت میکنند هر چه تعداد دندریتهای یک نورون بیشتر باشد قادر به دریافت اطلاعات زیادتری خواهد بود. در سطح خارجی برخی از دندریتها برآمدگیهای ریزی به نام خارهای دندریتی دیده میشود. تارهای عصبی که پیامهای حسهای پیکری را از گیرنده ها به نخاع می آورند در اصل دندریت هستند؛ ولی این تارها نسبتاً طویل اند و اغلب به وسیله غلاف میلین پوشیده میشوند و از نظر ظاهری شبیه به آکسونها هستند.
ساختمان و فیزیولوژی غشای نورون
غشای سیتوپلاسمی که اطراف جسم سلولی نورون و تارهای عصبی را فرامی گیرد از نظر ساختمانی مانند سایر غشاهای زنده سیتوپلاسمی در حدود ۸۰ آنگستروم ضخامت دارد و از ترکیبات فسفولیپید چربی فسفردار و پروتئین و کمی کربوهیدرات ساخته شده است و دارای آنزیمهای فراوان است کانالهای غشایی از جنس پروتئین بخش عمده ای از تبادلات سلول عصبی را با محیط خارج آن تأمین میکنند چربیهای غشای نورون به صورت عایقی عمل میکنند که بارهای مثبت و منفی را در سطوح خارجی و داخلی غشا از یکدیگر جدا میسازند آب و دی اکسید کربن و اوره و برخی مواد دیگر از غشای نورون عبور میکنند یونهای پتاسیم، سدیم و کلر نیز تحت شرایط خاصی از کانالهای غشای نورون میگذرند. ساختمان غشا جامد و بدون تغییر نیست و مولکولهای آن میتوانند جابه جا شوند و مواد دیگر را نیز حرکت دهند.
پتانسیل غشای عصبی (پتانسیل آرامش)
غشای عصبی در حالت آرامش یعنی در فقدان عامل محرک خارجی، قطبی (پولاریزه) است؛ یعنی بخش درونی غشا نسبت به سطح خارجی آن دارای مقداری بار منفی بوده و اختلاف سطح الکتریکی بین درون و بیرون غشا برقرار است. پتانسیل غشا در سایر یاخته های زنده نیز به میزان جزئی وجود دارد؛ ولی مقدار این پتانسیل در سلولهای عصبی و عضلانی از سایر سلولها زیادتر است و تحریک پذیر بودن این غشاها به پدیده های الکتریکی نسبتاً شدید آنها مربوط است. میزان پتانسیل آرامش غشا در تارهای عصبی در حدود ۷۰ و در تارهای عضلانی تقریباً ۹۰ میلی ولت است. با وارد کردن یک میکروالکترود به درون تار عصبی و اتصال آن به اسیلوسکوپ می توان اختلاف سطح الکتریکی بین این ناحیه و نواحی خارجی تار عصبی را اندازه گیری کرد. در این حالت گفته میشود که پتانسیل آرامش غشا ۷۰ میلی ولت است.
چگونگی ایجاد پتانسیل آرامش غشای عصبی
پتانسیل آرامش غشا زمانی به وجود می آید که تعداد یونهای مثبت در سطح بیرونی غشا کمی بیشتر از یونهای منفی و تعداد یونهای منفی در درون غشا کمی بیشتر از یونهای مثبت باشد. در ایجاد این نابرابری یونها چند عامل مؤثرند که مهمترین آنها یکی پمپ سدیم - پتاسیم و دیگری تفاوت نفوذ پذیری غشا نسبت به یونهای سدیم و پتاسیم در حالت آرامش است. در حالت آرامش نفوذ پذیری غشای سلول عصبی نسبت به یونهای پتاسیم در حدود صد برابر بیش از سدیم است. تعداد زیادی از ترکیبات درون و بیرون سلولها و تارهای عصبی به صورت یون و دارای بارهای الکتریکی مثبت یا منفی هستند یونهای سدیم و کلر مهم ترین یونهای محیط خارجی و یونهای پتاسیم پروتئینها و مولکولهای آلی فسفردار مهم ترین مواد محیط داخلی تارهای عصبی اند به طور کلی جابه جایی یونهای مثبت و منفی و نابرابری آنها در دو طرف غشا عامل ایجاد پتانسیل بیوالکتریک است و ترکیبات چربی غشا به صورت یک عایق در بین بارهای مثبت و منفی قرار دارد.
پمپ سدیم - پتاسیم
در غشای سیتوپلاسمی همه سلولهای بدن، پدیده انتقال فعال یونهای سدیم و پتاسیم وجود دارد که پمپ سدیم - پتاسیم نامیده می شود. این پمپ در غشای تارهای عصبی اهمیت بیشتری دارد یک مجموعه از آنزیمهای پروتئینی با صرف انرژی یونهای سدیم را از درون به بیرون و یونهای پتاسیم را از بیرون به درون سلول منتقل میکنند. این پروتئینهای غشایی که جایگاهی برای اتصال یونهای سدیم و پتاسیم دارند با صرف انرژی و به صورت هم زمان سه یون سدیم را با دو یون پتاسیم معاوضه میکنند یونهای سدیم و پتاسیم هر دو بار مثبت دارند؛ ولی با توجه به آنکه سه یون مثبت به خارج منتقل میشود و در مقابل آن فقط دو یون مثبت به درون میرود پمپ سدیم - پتاسیم تعادل بین تعداد یونهای مثبت و منفی را در بیرون و درون نورون بر هم میزند و باعث افزایش یونهای مثبت خارج نسبت به داخل میشود به همین دلیل پمپ سدیم - پتاسیم را پمپ الکتروژنیک تولید کننده پتانسیل الکتریکی مینامند مقدار پتانسیل الکتریکی غشا که مستقیماً بر اثر پمپ سدیم - پتاسیم ایجاد میشود از چند هزارم ولت بیشتر نیست و بخش بیشتری از پتانسیل آرامش غشای نورون نتیجه انتشار یونهای پتاسیم به خارج است.
پمپ سدیم - پتاسیم موجب میشود تا همواره تراکم پتاسیم در درون تار عصبی بیش از محیط خارج آن مایع میان بافتی بدن و تراکم سدیم در محیط خارجی تار عصبی بیش از محیط درون آن باشد. از سوی دیگر غشای عصبی در حالت آرامش نسبت به انتشار و جابه جایی پتاسیم نسبتاً نفوذ پذیر و نسبت به سدیم تقریباً نفوذ ناپذیر است. بنابراین یونهای سدیم که با صرف انرژی به بیرون ریخته شده اند نمی توانند در جهت شیب تراکم خود به داخل تار عصبی وارد شوند؛ در حالی که تعدادی از یونهای پتاسیم داخلی پیوسته در جهت شیب تراکم خود به سطح خارجی غشا انتشار می یابند. انتشار یونهای پتاسیم به خارج اختلاف سطح الکتریکی غشا را تا حدود ۷۰ میلی ولت افزایش میدهد بنابراین بخش اعظم پتانسیل آرامش غشا با انتشار یونهای پتاسیم به خارج غشا به وجود می آید؛ ولی این انتشار خود حاصل کار پمپ سدیم - پتاسیم است که اختلاف تراکم این دو یون را در دو طرف غشا ایجاد کرده است.
در واقع در حالت استراحت فقط خروج تعداد بسیار کمی از یونهای پتاسیم در حدود یک یون از هر ۱۰۰ هزار یون داخلی برای ایجاد پتانسیل آرامش غشا کافی است؛ زیرا با خروج پتاسیم و ایجاد پتانسیل غشا یک نیروی الکتریکی در راستای پیشگیری از خروج پتاسیم بیشتر وارد عمل میشود. در این حالت بین نیروی انتشار که پتاسیم را به بیرون میراند و دافعه یونهای مثبت که مانع خروج یونهای مثبت پتاسیم میشود تعادل برقرار می گردد؛ از این روست که پتانسیل آرامش غشا را پتانسیل تعادل نیز نام گذاری کرده اند. در علم بیوفیزیک قوانین و فرمولهایی وجود دارد که به کمک آنها می توان با دانستن اختلاف تراکم یونهای مثبت و منفی موجود در دو طرف غشا و توانایی انتشار این یونها از غشا مقدار اختلاف سطح الکتریکی ایجاد شده را محاسبه کرد. اگر با توجه به اختلاف تراکم یونهای پتاسیم در دو محیط درونی و بیرونی تارهای عصبی، میزان پتانسیل غشای حاصل از انتشار این یون را محاسبه کنیم رقمی در حدود ۹۰ هزارم ولت یعنی کمی بیش از مقدار واقعی پتانسیل آرامش غشای نورون به دست می آید. علت این تفاوت این است که غشای عصبی در حالت آرامش نسبت به سایر یونها کاملاً نفوذ ناپذیر نیست و تعداد کمی یون سدیم به درون تارهای عصبی انتشار یافته از میزان پتانسیل آرامش می کاهد. ایجاد پتانسیل آرامش غشای عصبی امکان تحریک سریع آن را فراهم می سازد. در فیزیولوژی اعصاب با توجه به اینکه پتانسیل آرامش را با وارد کردن میکروالکترود به درون تار بررسی و پتانسیل درون را نسبت به بیرون اندازه گیری کنند این پتانسیل را به صورت منفی ۷۰ تا ۹۰ نشان میدهند.
پتانسیل عمل
هنگامی که بخشی از غشای نورون تحریک میشود یک دگرگونی بیوالکتریکی به نام پتانسیل عمل در غشای آن پدید می آید که از محل تحریک به نواحی دیگر، در طول غشای تار عصبی انتشار می یابد. این پدیده موج یا جریان عصبی است. فقط غشای سلولهای عصبی و عضلانی میتوانند پتانسیل عمل تولید کنند؛ به همین دلیل غشای این دو نوع سلول تحریک پذیر خوانده میشوند محرکها انواع مختلفی دارند و ممکن است مکانیکی شیمیایی حرارتی نوری و یا الکتریکی باشند. در بررسیهای تجربی برای تحریک تار عصبی معمولاً از محرکهای الکتریکی استفاده می کنند؛ زیرا شدت این محرکها به دقت قابل تغییر و تنظیم است و اثر آنها به سرعت ظاهر و قطع میشود و در محدوده مناسب آسیبی به غشای عصبی وارد نمی کند.
مراحل پتانسیل عمل
پس از تأثیر محرک در غشای تحریک پذیر، پتانسیل آرامش آن ناحیه ابتدا به آرامی و سپس به سرعت کاهش می یابد و به صفر می رسد و حتى درون غشا نسبت به سطح خارجی آن بار مثبت پیدا می کند. این تغییرات با وارد کردن یک میکروالکترود به درون تار و اتصال آن به دستگاهی به نام نوسان نگار اسیلوسکوپ) به صورت یک خط بالارو نمایان میشود که مرحله دپولاریزاسیون از بین رفتن پولاریزاسیون خوانده میشود. در قله دپولاریزاسیون تار عصبی درون تار نسبت به بیرون آن در حدود ۳۰ تا ۴۰ هزارم ولت بار مثبت پیدا کرده است. پس از آن شاخه پایین رونده منحنی ثبت میشود که نشانه بازگشت مجدد پتانسیل آرامش یا رپولاریزاسیون غشاست. در پایان این مرحله در مدت کوتاهی میزان پتانسیل آرامش تار عصبی حتی از حالت عادی آن نیز کمی بیشتر می شود که به این بخش مرحله هیپرپولاریزاسیون می گویند. سرانجام پتانسیل آرامش غشا در حد عادی خود قرار میگیرد. زمان این مراحل به ویژگیهای فیزیولوژیکی و قطر تارها بستگی دارد؛ ولی به طور کلی مجموعه آنها بیش از چند هزارم ثانیه طول نمی کشد.
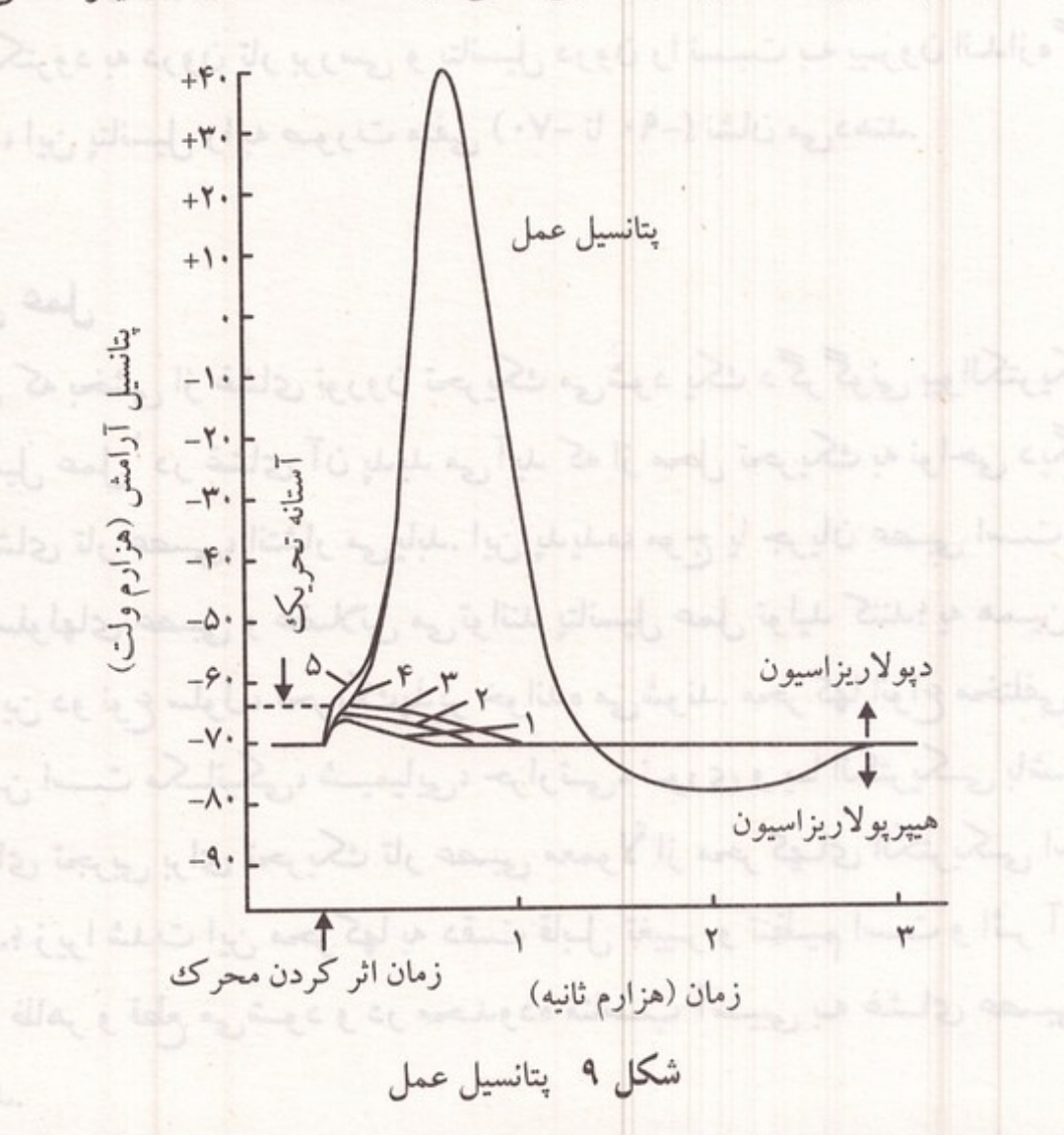
شکل ۹: پتانسیل عمل، نمایش مراحل دپولاریزاسیون، رپولاریزاسیون و هیپرپولاریزاسیون.
پتانسیل موضعی (محلی)
اگر یک محرک ضعیف که قادر به ایجاد پتانسیل عمل واقعی نباشد بر تار عصبی وارد کنیم یک پاسخ موضعی ایجاد می شود که اثر آن هر چه از محل تحریک دورتر شویم کمتر میشود معمولاً در یک تار عصبی پتانسیل موضعی و پتانسیل عمل منتشر شونده به یکدیگر وابسته اند؛ به این ترتیب که تا زمانی که شدت محرک به حد معینی نرسیده باشد پتانسیل موضعی به وجود می آید و پس از آن با رسیدن شدت محرک به حد لازم پتانسیل عمل منتشر شونده به جریان می افتد.
اساس یونی پتانسیل عمل
غشای عصبی در حالت استراحت نسبت به یونهای سدیم که تعداد آنها در بیرون بسیار بیشتر از درون نورون است، تقریباً نفوذ ناپذیر است و بنابر اصطلاح دریچه های سدیم در غشا بسته اند. اثر محرک در غشا باعث افزایش نفوذ پذیری آن نسبت به سدیم میشود. ورود یونهای مثبت که بار مثبت دارند وضعیت الکتریکی غشا را تغییر میدهد و آن را دپولاریزه میکند تا زمانی که اثر محرک در غشا میزان پتانسیل آرامش را از حدود ۷۰ میلی ولت به رقم تقریبی ۵۵ میلی ولت نرساند، پتانسیل عمل منتشر شونده ایجاد نمیشود و اثر محرک به صورت پتانسیل موضعی باقی میماند اگر شدت محرک به حد لازم برسد دریچه های سدیم به تعداد کافی باز میشود و ورود یونهای سدیم که در جهت شیب تراکم آنها صورت می گیرد باعث دپولاریزاسیون غشا میگردد. باز بودن دریچه های سدیم بسیار زودگذر است و این دریچه ها در قله پتانسیل عمل دوباره بسته میشوند و دپولاریزاسیون تار فراتر نمی رود.
بسته شدن دریچه های سدیم و افزایش نفوذپذیری غشا نسبت به یونهای پتاسیم که از این لحظه به بعد رخ میدهد باعث بازگشت مجدد پتانسیل آرامش به غشای عصبی میشود در مرحله دپولاریزاسیون تراوایی غشا نسبت به یونهای پتاسیم تغییر نمی کند؛ ولی در این مرحله میزان افزایش بارهای مثبت درون غشا که از ورود سدیم ناشی میشود بیش از مقدار کاهش آنها بر اثر خروج پتاسیم است. برای ایجاد پتانسیل عمل ورود تعداد کمی یون سدیم کافی است و سدیم وارد شده به وسیله پمپ سدیم با صرف مقدار کمی انرژی دوباره به بیرون رانده می شود. در هنگام تحریک غشای عصبی باز شدن دریچه های سدیم شبیه کار کرد یک بازخورد مثبت است؛ به این ترتیب که اثر تحریک ابتدا باعث نشت مختصر یونهای سدیم به درون میشود و پتانسیل غشا را کاهش میدهد و کاهش پتانسیل غشا به باز شدن دریچه های سدیم بیشتر و دپولاریزه شدن غشا منجر میشود؛ از این رو گفته می شود که باز و بسته شدن دریچه های سدیم در غشاهای تحریک پذیر به ولتاژ غشا وابسته است.
عمل مواد بی حس کننده موضعی مانند نووکائین و گزیلو کائین به جلوگیری از ورود یونهای سدیم به تار عصبی مربوط است. این مواد با پیشگیری از ورود سدیم مانع ایجاد پتانسیل عمل میشوند کوکائین که از گیاه کوکا به دست می آید اولین ماده ای بود که به عنوان بی حس کننده موضعی شناخته شد ولی به علت خواص اعتیاد آوری که دارد ممنوع گردید و بی حس کننده های موضعی صناعی مانند پروکائین ساخته شوند. با استفاده از این مواد در اعصاب محیطی یا مرکزی بی حسی نخاعی پا اپی دورال ایجاد و امکان جراحی بدون احساس درد را فراهم میکنند. بعضی مواد دیگر مانند حشره کشها با از کار انداختن پمپ سدیم باعث اختلال در تحریک اعصاب می شوند. کاهش کلسیم خون باعث میشود تا دریچه های سدیم غشای عصبی در حالت آرامش به خوبی بسته نشوند و ورود یونهای سدیم بیشتر به درون تار عصبی موجب میشود تا بدون وجود محرک امواج عصبی درون زا تولید شود.
آستانه تحریک و قانون همه یا هیچ
کمترین شدت محرکی که بتواند غشای عصبی را تحریک و پتانسیل عمل در آن ایجاد کند آستانه تحریک خوانده می شود. در این حالت کاهش پتانسیل آرامش تار عصبی به حدی رسیده است که دریچه های سدیم وابسته به ولتاژ باز شوند و پتانسیل عمل را به راه اندازند. این حد از پتانسیل آرامش را سطح آتش یا سطح شلیک پتانسیل عمل مینامند محرکهای ضعیف تر از آستانه تحریک چنان که قبلاً گفته شده است فقط یک دپولاریزاسیون محلی ایجاد می کنند که اثر آن در فاصله کوتاهی از بین میرود بر اساس قانون همه یا هیچ که در یک تار عصبی صدق میکند پاسخ تار عصبی به محرکهای شدیدتر از آستانه تحریک از نظر شدت دپولاریزاسیون یعنی ولتاژ پتانسیل عمل با پاسخی که با شدت آستانه تولید میشود یکسان است و این شدت در طول تار عصبی نیز ثابت می ماند. به موجب قانون همه یا هیچ هرگاه پتانسیل آرامش غشای عصبی بر اثر یک محرک به میزان معینی کاهش یافت پتانسیل عمل یا پاسخ منتشر شونده موج عصبی در آن به وجود می آید که ولتاژ آن به حداکثر ممکن است و با افزایش شدت محرک تغییر نمی کند. آنچه در دستگاه عصبی باعث تمیز و تشخیص محرکهای شدید و ضعیف می شود این است که از یک سو با افزایش شدت محرک تعداد بیشتری از تارهای عصبی تحریک میشوند و از سوی دیگر در هر تار نیز با افزایش شدت محرک تا حد معینی بر تعداد پتانسیل عملهای ایجاد شده افزوده میشود بدون آنکه شدت (ولتاژ) هر پتانسیل عمل تغییر کند. در یک عصب که خود از تعدادی تار عصبی ساخته شده است قانون همه یا هیچ صادق نیست؛ زیرا با افزایش شدت محرک به تدریج تعداد بیشتری از تارهای عصبی تحریک میشوند و پاسخ کلی ثبت شده شدت بیشتری می یابد.
شدت پایه و زمان مفید
اگر شدت آستانه تحریک را برای محرکهایی اندازه گیری کنیم که زمان اثر کردن آنها را به تدریج کاهش میدهیم، خواهیم دید که بین شدت محرک و مدت آن رابطه معکوس وجود دارد یعنی از حدی به بعد با کاهش زمان محرک شدت آستانه تحریک افزایش می یابد و در صورتی که زمان محرک از حدی بیشتر شود آستانه تحریک به حداقل خود می رسد و از آن پس ثابت می ماند. این مدت محرک را که در آن شدت آستانه به کمترین میزان خود می رسد زمان مفید نامیده اند.
مراحل تحریک ناپذیری
یکی از ویژگیهای عملی غشاهای تحریک پذیر یاخته های عصبی این است که نمیتوانند بلافاصله پس از ایجاد یک پتانسیل عمل دوباره تحریک شوند لذا برای مدت کوتاهی تحریک ناپذیر می مانند. این دوره خود شامل دو مرحله تحریک ناپذیری مطلق و نسبی است. در مرحله اول که ۱ تا ۲ هزارم ثانیه طول میکشد آستانه تحریک تار عصبی بی نهایت و تحریک پذیری آن صفر است. این زمان که مرحله تحریک ناپذیری مطلق است برای بازگشت شرایط طبیعی غشای نورون ضرورت دارد. پس از مرحله مذکور غشای عصبی به تدریج تحریک پذیری طبیعی خود را باز می یابد و آستانه تحریک آن تا حد معمولی پایین می آید. این مرحله که تحریک ناپذیری نسبی خوانده میشود طولانی تر از مرحله قبلی است و ۱۰ تا ۱۵ هزارم ثانیه طول میکشد در این مرحله می توان به کمک محرکهای شدیدتر از آستانه تحریک معمولی تار عصبی را تحریک کرد. مرحله تحریک ناپذیری مطلق هر نورون به طور تقریبی با دپولاریزاسیون آن مقارن است و مرحله تحریک ناپذیری نسبی تا پس از شاخه رپولاریزاسیون ادامه دارد. وجود مراحل تحریک ناپذیری باعث محدود شدن تعداد پتانسیلهای عملی میشود که یک تار میتواند در هر ثانیه تولید کند اعصاب پستانداران میتواند صدها پتانسیل عمل در ثانیه ایجاد کند و توانایی تولید پتانسیل عمل در برخی از تارهای عصبی به بیش از هزار در ثانیه می رسد.
زمان نهفته
از لحظه ای که محرک بر تار عصبی وارد می شود تا لحظه ای که پتانسیل عمل آغاز میگردد زمان نهفته خوانده میشود این مدت در تارهای عصبی مختلف متفاوت است؛ هر چه تحریک پذیری تار عصبی بیشتر باشد زمان نهفته آن کوتاه تر است. زمان نهفته در تارهای عصبی مختلف از ۱ تا دو ۲ هزارم ثانیه طول می کشد.
هدایت موج عصبی
پتانسیل عملی که در محل تحریک به وجود می آید به با شدت یکسان و بدون کاهش در طول تار عصبی سیر میکند پتانسیل عمل همواره منطقه معینی را اشغال میکند که طول موج آن است. هر منطقه تحریک شده باعث تحریک ناحیه مجاور میشود و خود به حالت آرامش باز می گردد. پتانسیل عمل به صورت یک ناحیه دپولاریزه شده حلقه مانند از یک سر آکسون به جانب دیگر آن حرکت می کند. باید توجه داشت که هدایت پتانسیل عمل در طول تار عصبی یک پدیده انفعالی نیست و تار عصبی مانند یک هادی ساده که جریان الکتریکی عصبی را در طول خود هدایت میکند نیست بلکه پتانسیل عملی که در یک ناحیه به وجود آمده است برای نواحی مجاور خود به صورت یک محرک الکتریکی عمل می کند و آن نواحی را که در حالت آرامشاند دپولاریزه می کند و باعث ایجاد همان تغییرات یونی لازم برای ایجاد پتانسیل عمل میشود؛ این پدیده در تمام طول تار عصبی تکرار میگردد. به همین دلیل است که موج عصبی در هدایت خود به نواحی دوردست کاهش نمی یابد و شدت آن در ابتدا و انتهای تار عصبی یکسان است. معمولاً هدایت پتانسیل عمل را در طول تار عصبی به یک طناب آغشته به باروت تشبیه میکنند که اگر یک سر آن آتش زده شود شعله به صورت یکنواخت و با شدت برابر در طول طناب پیش میرود انتشار موج عصبی در طول یک تار عصبی که به صورت تجربی در خارج از بدن مورد بررسی قرار گیرد در هر دو جهت امکان پذیر است و اگر در وسط یک تار عصبی، محرکی را اثر دهیم دو پتانسیل عمل به وجود می آید که در جهت عکس یکدیگر به سوی دو انتهای تار سیر خواهند کرد در بدن با توجه به اینکه در حالت عادی پتانسیل عمل فقط در یک سر تار عصبی به وجود می آید مسیر آن یک طرفی است. ضمناً ساختمان ویژه سیناپسها یعنی محل ارتباط نورونها با یکدیگر به صورتی است که موج عصبی را فقط در یک جهت عبور می دهد.
سرعت هدایت موج عصبی
سرعت هدایت موج عصبی در طول یک تار ثابت ولی در تارهای مختلف متفاوت است. هر چه قطر تار عصبی بیشتر باشد سرعت هدایت موج عصبی در آن زیادتر است وجود میلین نیز باعث افزایش سرعت انتشار موج عصبی میشود. سرعت هدایت پتانسیل عمل در قطورترین تارهای میلین دار انسان در حدود ۱۲۰ متر در ثانیه و در نازک ترین تارهای بدون ميلين تقریباً ۰/۵ متر در ثانیه است. گرم و سرد کردن تار عصبی باعث افزایش و کاهش سرعت موج عصبی میشود. اگر شدت سرما از حدی بیشتر شود انتشار موج عصبی متوقف میشود. در هنگام تب سرعت سیر امواج عصبی در بدن افزایش می یابد. تارهای عصبی را با توجه به قطر آنها وجود یا فقدان میلین در آنها و قطر میلین به سه دسته B A و C تقسیم میکنند. دو گروه A و B دارای میلین و گروه C بدون میلین هستند تارهای گروه A که قطورترین تارهای عصبی را تشکیل میدهند به چهار زیر گروه آلفا بتا گاما و دلتا تقسیم میشوند. تارهای زیر گروه آلفا قطور و میلین دار هستند و فاصله بین گرههای رانویه در آنها زیاد است. سرعت سیر موج عصبی در این تارها بیشترین مقدار را دارد زمان نهفته و مراحل تحریک ناپذیری آنها کوتاه است. در سایر زیر گروهها سرعت به ترتیب کمتر است. تارهای گروه B میلین کمتری دارند و سرعت جریان عصبی در آنها کمتر از گروه A و بین ۳ تا ۱۵ متر در ثانیه است. گروه C نازک ترین تارهای عصبی بدون میلین را شامل میشود که نسبت به سایر تارها سرعت سیر کمتر و آستانه تحریک بالاتری دارند. سرعت هدایت موج عصبی در این تارها بین ۰/۵ تا ۲ متر در ثانیه است.
هدایت جهشی
وجود غلاف میلین در تارهای عصبی که به صورت یک لایه عایق غشای آکسون را میپوشاند باعث میشود تا فقط در گرههای را نویه پتانسیل آرامش تولید شود و در نتیجه فقط این گرهها قادر به ایجاد پتانسیل عمل باشند. به همین دلیل هدایت موج عصبی در تارهای میلین دار به صورت جهشی صورت می گیرد؛ یعنی از یک گره به گره بعدی منتقل میشود؛ این نوع انتشار باعث افزایش سرعت موج عصبی و صرفه جویی در مصرف انرژی میگردد. وجود مایع میان بافتی در سطح خارجی تارهای عصبی که هادی جریان الکتریکی است، مدار خارجی انتقال تحریک از یک گره به گره بعدی را امکان پذیر میکند. بخش داخلی مدار تحریک این گرهها به وسیله اکسوپلاسم درون تار تأمین می شود. در تارهای عصبی میلین دار فقط گرههای را نویه میتوانند یونها را به میزان لازم برای ایجاد پدیده های الکتریکی از خود عبور دهند و میزان عبور یونها به علت کوچک بودن مساحت گرههای را نویه بسیار کمتر از تارهای عصبی بدون میلین است.
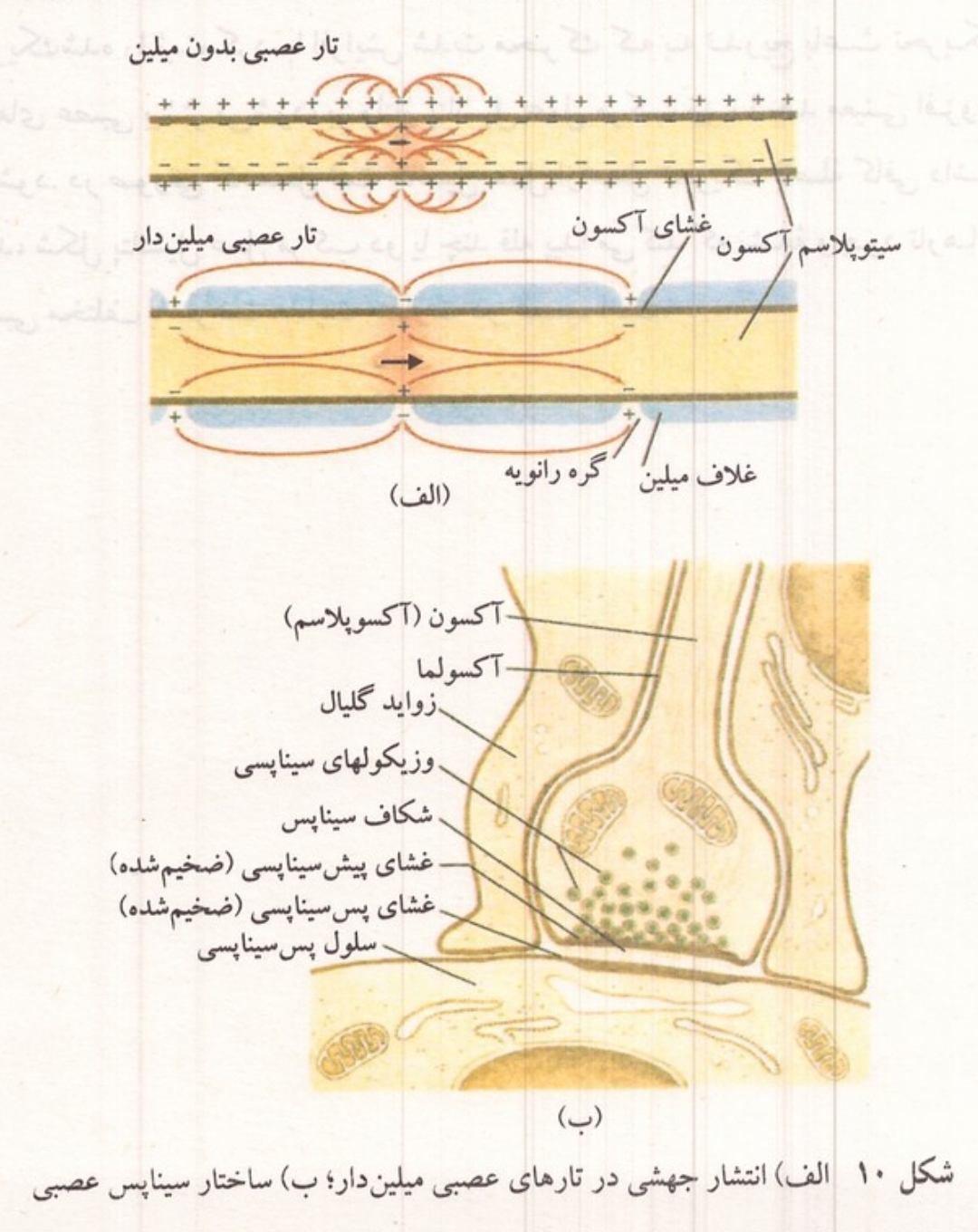
شکل ۱۰: (الف) انتشار جهشی در تارهای عصبی میلین دار (ب) ساختار سیناپس عصبی
ساخته نشدن میلین و یا تحلیل رفتن آن باعث بیماریهای عصبی مهمی میشود که ام اس (MS) یا مالتیپل اسکلروزیس یکی از آنهاست. این بیماری که از نوع خود ایمنی است معمولاً بین سنین ۲۰ تا ۵۰ سالگی بروز می کند و در زنان شایع تر از مردان است. در این بیماری پادتنها و گلبولهای سفید خون به غلاف میلین حمله می کنند و باعث التهاب و تحلیل رفتن تدریجی غلاف میلین و اختلال در هدایت امواج عصبی میشوند. از نشانه های اولیه این بیماری ضعف پاها، التهاب عصب بینایی و اختلال در تکلم و بلع است.
پتانسیل عمل مرکب
هر عصب یا تنه عصبی از تعدادی تار عصبی ساخته شده است. با قرار دادن الکترودهای خارجی بر روی عصب و اتصال آنها به اسیلوسکوپ و تحریک عصب با محرک نسبتاً قوی میتوان پتانسیل عمل مجموعه تارهای تحریک شده را ثبت کرد با افزایش شدت محرک که به تدریج باعث تحریک تارهای عصبی بیشتر میشود بر ولتاژ پتانسیل عمل مرکب نیز تا حد معینی افزوده می شود. در صورتی که محل ثبت پتانسیل عمل از محل تحریک فاصله کافی داشته باشد شکل پتانسیل عمل مرکب دو یا چند قله پیدا میکند که نشانه وجود تارهای عصبی مختلف با سرعت هدایت متفاوت در عصب است.
رسانههای تکمیلی فصل
پادکست
ویدیو
آزمونک فصل سوم (سوالات تشریحی)
دانش خود را بر اساس مطالب این فصل بسنجید.
۱. «نورون»، «تار عصبی» و «عصب» را تعریف کنید و تفاوت آنها را بیان کنید.
نمایش پاسخ
پاسخ: نورون، واحد ساختمانی و عملی دستگاه عصبی است (کل سلول). تار عصبی، به هر یک از رشتههای سیتوپلاسمی (آکسون یا دندریت) که از جسم سلولی خارج میشوند، گفته میشود. عصب، اجتماعی از تعدادی تار عصبی است که توسط غلافی پیوندی پوشیده شدهاند.
۲. سیتوپلاسم سلول عصبی (نوروپلاسم) حاوی چه اندامکهای مهمی است؟ (حداقل ۴ مورد)
نمایش پاسخ
پاسخ: میتوکندریهای فراوان، شبکه آندوپلاسمی و ریبوزومها (که اجسام نیسل را میسازند)، دستگاه گلژی، لیزوزومها، نوروفیبریلها و نوروتوبولها.
۳. «اجسام نیسل» چه هستند و چه نقشی دارند؟
نمایش پاسخ
پاسخ: دانههای درشتی هستند که از تجمع دانه های ریبوزوم و شبکه آندوپلاسمی ساخته شدهاند. این اجسام رنگهای قلیایی را جذب میکنند و در پروتئینسازی و انتقال آنها نقش اساسی دارند.
۴. نقش نوروفیبریلها و نوروتوبولها در نورون چیست؟
نمایش پاسخ
پاسخ: این رشتهها و لولههای باریک، نوعی اسکلت سیتوپلاسمی تشکیل میدهند که به حفظ شکل نورون و تارهای عصبی کمک میکنند.
۵. نورونها از نظر طرز خارج شدن تارها از جسم سلولی به چند گروه تقسیم میشوند؟ نام ببرید و برای هر کدام مثالی بزنید.
نمایش پاسخ
پاسخ: به سه گروه تقسیم میشوند:
۱. یک قطبی: آکسون و دندریت از یک ناحیه خارج میشوند (مانند نورونهای عقدههای نخاعی انسان).
۲. دو قطبی: آکسون از یک ناحیه و دندریتها از ناحیه دیگر خارج میشوند (مانند گیرندههای بویایی و برخی نورونهای شبکیه).
۳. چند قطبی: آکسون از یک ناحیه و دندریتها از چند ناحیه دیگر خارج میشوند (مانند نورونهای حرکتی نخاع، سلولهای پورکینیه مخچه).
۶. «میلین» چیست؟ از چه موادی تشکیل شده و چه تاثیری بر رنگ بافت عصبی دارد؟
نمایش پاسخ
پاسخ: میلین یک ماده پروتئینی و چربی فسفردار (فسفولیپید) سفیدرنگ است که برخی آکسونها را به صورت غلافی ناپیوسته میپوشاند و باعث رنگ سفید اعصاب و نواحی سفید مغز و نخاع میشود.
۷. «گرههای رانویه» (Nodes of Ranvier) چه هستند و در کجا قرار دارند؟
نمایش پاسخ
پاسخ: گرههای رانویه، گسستگیهایی در غلاف میلین هستند که به فواصل مساوی در طول آکسونهای میلیندار دیده میشوند. در این نواحی، غشای آکسون (آکسولم) فاقد پوشش میلین است.
۸. سلولهای سازنده میلین در «اعصاب پیرامونی» (PNS) و «مراکز عصبی» (CNS) چه نام دارند؟
نمایش پاسخ
پاسخ: در اعصاب پیرامونی توسط سلولهای شوان و در مراکز عصبی (مغز و نخاع) توسط اولیگودندروگلیا ساخته میشود.
۹. تفاوت اصلی ساختاری دندریتها با آکسونها چیست؟
نمایش پاسخ
پاسخ: دندریتها معمولاً انشعابات زیادی دارند و کوتاهترند (هرچند استثنا وجود دارد)، قطر قاعده آنها بیشتر از انتهایشان است و حاوی اندامکهایی مانند ریبوزوم و شبکه آندوپلاسمی (اجسام نیسل) هستند. آکسون یک رشته منفرد است که میلین دارد و اندامکهای سنتزی در آن کمتر است.
۱۰. «خارهای دندریتی» (Dendritic Spines) چه هستند؟
نمایش پاسخ
پاسخ: برآمدگیهای ریزی در سطح خارجی برخی از دندریتها هستند که محل دریافت اطلاعات (سیناپس) از نورونهای دیگر میباشند.
۱۱. غشای نورون از چه ترکیباتی ساخته شده و نقش چربیهای آن چیست؟
نمایش پاسخ
پاسخ: از ترکیبات فسفولیپید (چربی فسفردار) و پروتئین و کمی کربوهیدرات ساخته شده است. چربیهای غشا به صورت عایق عمل میکنند و بارهای مثبت و منفی را در دو سطح غشا از هم جدا میسازند.
۱۲. «پتانسیل آرامش» (Resting Potential) چیست و مقدار آن در تار عصبی چقدر است؟
نمایش پاسخ
پاسخ: در حالت آرامش (نبود تحریک)، غشای عصبی قطبی (پولاریزه) است؛ یعنی بخش درونی غشا نسبت به بیرون، بار منفی دارد. این اختلاف سطح الکتریکی، پتانسیل آرامش نام دارد و میزان آن در تارهای عصبی حدود ۷۰- میلی ولت است.
۱۳. دو عامل اصلی و مؤثر در ایجاد پتانسیل آرامش غشا کدامند؟
نمایش پاسخ
پاسخ: ۱. پمپ سدیم-پتاسیم (که شیب غلظت را ایجاد میکند).
۲. تفاوت نفوذپذیری غشا نسبت به یونها در حالت آرامش (نفوذپذیری بسیار زیاد به پتاسیم و بسیار کم به سدیم).
۱۴. توزیع یونهای سدیم (Na+), پتاسیم (K+) و پروتئینهای آنیونی در حالت آرامش چگونه است؟
نمایش پاسخ
پاسخ: در محیط خارجی (بیرون سلول): سدیم و کلر زیاد است.
در محیط داخلی (درون سلول): پتاسیم، پروتئینها و مولکولهای آلی فسفردار (که بار منفی دارند) زیاد است.
۱۵. مکانیسم عمل «پمپ سدیم-پتاسیم» را به طور دقیق شرح دهید (چه تعداد یون به کجا منتقل میشود).
نمایش پاسخ
پاسخ: این پمپ مجموعهای از آنزیمهای پروتئینی در غشا است که با صرف انرژی (ATP)، به صورت همزمان سه (۳) یون سدیم (Na+) را از درون به بیرون سلول و دو (۲) یون پتاسیم (K+) را از بیرون به درون سلول منتقل میکند.
۱۶. چرا پمپ سدیم-پتاسیم را «الکتروژنیک» (تولید کننده پتانسیل) مینامند؟
نمایش پاسخ
پاسخ: زیرا با انتقال غیرمساوی بارهای مثبت (۳ مثبت به خارج در ازای ۲ مثبت به داخل)، به طور مستقیم باعث افزایش یونهای مثبت در خارج نسبت به داخل شده و بخش کوچکی از پتانسیل منفی غشا را ایجاد میکند.
۱۷. نقش اصلی پمپ سدیم-پتاسیم در ایجاد پتانسیل آرامش چیست؟ (نقش مستقیم در مقابل نقش غیرمستقیم)
نمایش پاسخ
پاسخ: نقش مستقیم آن (اثر الکتروژنیک) ناچیز است. نقش اصلی آن ایجاد و حفظ اختلاف تراکم (شیب غلظت) یونهای سدیم و پتاسیم در دو طرف غشا است. بخش اعظم پتانسیل آرامش (۷۰- میلی ولت) نتیجه انتشار (نشت) یونهای پتاسیم به خارج از سلول (در جهت شیب تراکمی که پمپ ایجاد کرده) میباشد.
۱۸. «پتانسیل عمل» (Action Potential) چیست و در چه سلولهایی رخ میدهد؟
نمایش پاسخ
پاسخ: یک دگرگونی بیوالکتریکی سریع در غشای نورون است که در پاسخ به تحریک ایجاد شده و در طول تار عصبی انتشار مییابد (موج یا جریان عصبی). این پدیده فقط در غشای سلولهای عصبی و عضلانی (سلولهای تحریکپذیر) رخ میدهد.
۱۹. مراحل پتانسیل عمل را به ترتیب نام ببرید و توضیح دهید در هر مرحله پتانسیل غشا چه تغییری میکند.
نمایش پاسخ
پاسخ: ۱. دپولاریزاسیون (Depolarization): پتانسیل آرامش (۷۰-) به سرعت کاهش یافته، به صفر رسیده و حتی مثبت میشود (تا حدود ۳۰+ یا ۴۰+ میلی ولت).
۲. رپولاریزاسیون (Repolarization): بازگشت سریع پتانسیل غشا از حالت مثبت به حالت منفی (پتانسیل آرامش).
۳. هیپرپولاریزاسیون (Hyperpolarization): در مدت کوتاهی، پتانسیل غشا حتی از حالت آرامش عادی نیز منفیتر میشود.
۲۰. تفاوت «پتانسیل موضعی» (محلی) و «پتانسیل عمل» چیست؟
نمایش پاسخ
پاسخ: پتانسیل موضعی در اثر محرک ضعیف (زیر آستانه) ایجاد میشود، منتشر شونده نیست و اثر آن هر چه از محل تحریک دورتر شویم کمتر میشود. اما پتانسیل عمل در اثر محرک آستانه یا قویتر ایجاد شده و در تمام طول تار عصبی منتشر میشود.
۲۱. اساس یونی (علت یونی) مرحله «دپولاریزاسیون» پتانسیل عمل چیست؟
نمایش پاسخ
پاسخ: باز شدن سریع دریچههای سدیم (Na+) وابسته به ولتاژ و ورود (نفوذ) یونهای سدیم به درون تار عصبی در جهت شیب تراکم.
۲۲. اساس یونی (علت یونی) مرحله «رپولاریزاسیون» پتانسیل عمل چیست؟
نمایش پاسخ
پاسخ: بسته شدن دریچههای سدیم و افزایش نفوذپذیری غشا به یونهای پتاسیم (K+) (باز شدن کانالهای پتاسیمی وابسته به ولتاژ) و خروج یونهای پتاسیم از سلول.
۲۳. «سطح آتش» یا آستانه تحریک (Threshold) به چه معناست؟
نمایش پاسخ
پاسخ: کمترین شدت محرکی (یا سطحی از پتانسیل، حدود ۵۵- میلی ولت) که بتواند دریچههای سدیم وابسته به ولتاژ را به تعداد کافی باز کند تا پتانسیل عمل (موج منتشر شونده) ایجاد شود.
۲۴. مکانیسم عمل مواد بیحس کننده موضعی مانند نووکائین چیست؟
نمایش پاسخ
پاسخ: این مواد با جلوگیری از ورود یونهای سدیم به تار عصبی (مسدود کردن کانالهای سدیم)، مانع ایجاد پتانسیل عمل و در نتیجه انتقال پیام درد میشوند.
۲۵. کاهش کلسیم خون چه تأثیری بر تحریکپذیری اعصاب دارد و چرا؟
نمایش پاسخ
پاسخ: باعث افزایش تحریکپذیری اعصاب میشود. زیرا کاهش کلسیم موجب میشود دریچههای سدیم در حالت آرامش به خوبی بسته نشوند، یون سدیم به درون نشت کرده و امواج عصبی درونزا (خودبخودی) تولید شود.
۲۶. «قانون همه یا هیچ» (All-or-None Law) در یک تار عصبی را توضیح دهید.
نمایش پاسخ
پاسخ: این قانون میگوید اگر یک تار عصبی با محرکی ضعیفتر از آستانه تحریک شود، هیچ پاسخی نمیدهد (هیچ). اما اگر با محرک آستانه یا قویتر از آن تحریک شود، یک پتانسیل عمل با حداکثر ولتاژ ممکن ایجاد میکند (همه). ولتاژ پتانسیل عمل با افزایش شدت محرک، بیشتر نمیشود.
۲۷. اگر قانون «همه یا هیچ» برقرار است، دستگاه عصبی چگونه شدت محرکهای قوی و ضعیف را از هم تشخیص میدهد؟
نمایش پاسخ
پاسخ: از دو طریق:
۱. تعداد (فرکانس) پتانسیلهای عمل: محرک قویتر، تعداد بیشتری پتانسیل عمل در ثانیه در همان تار ایجاد میکند.
۲. تعداد تارهای عصبی: محرک قویتر، تعداد بیشتری از تارهای عصبی را تحریک میکند.
۲۸. «مرحله تحریک ناپذیری مطلق» (Absolute Refractory Period) چیست و همزمان با کدام بخش پتانسیل عمل است؟
نمایش پاسخ
پاسخ: مدتی کوتاه (۱ تا ۲ هزارم ثانیه) پس از شروع پتانسیل عمل است که در آن آستانه تحریک بینهایت بوده و تار عصبی تحت هیچ شرایطی (حتی با محرک قوی) دوباره تحریک نمیشود. این مرحله تقریباً مقارن با مرحله دپولاریزاسیون است.
۲۹. «مرحله تحریک ناپذیری نسبی» (Relative Refractory Period) چیست؟
نمایش پاسخ
پاسخ: مرحلهای پس از تحریکناپذیری مطلق است که در آن غشا به تدریج تحریکپذیری خود را باز مییابد. در این مرحله، میتوان تار عصبی را تحریک کرد، اما فقط به کمک محرکهایی شدیدتر از آستانه تحریک معمولی.
۳۰. هدایت موج عصبی (پتانسیل عمل) در طول تار عصبی چگونه رخ میدهد؟ (مکانیسم انتشار)
نمایش پاسخ
پاسخ: هدایت یک پدیده انفعالی (مانند جریان در سیم) نیست. بلکه پتانسیل عملی که در یک ناحیه به وجود آمده، برای ناحیه مجاور خود به صورت یک محرک الکتریکی عمل میکند و آن ناحیه را دپولاریزه کرده و باعث ایجاد پتانسیل عمل «جدید» در آن ناحیه میشود. این پدیده در تمام طول تار عصبی تکرار میگردد.
۳۱. چه عواملی بر سرعت هدایت موج عصبی تأثیر دارند؟ (حداقل ۳ مورد)
نمایش پاسخ
پاسخ: ۱. قطر تار عصبی: هر چه قطر بیشتر باشد، سرعت بیشتر است.
۲. وجود میلین: تارهای میلیندار سرعت بسیار بیشتری (هدایت جهشی) دارند.
۳. دما: گرما سرعت را افزایش و سرما آن را کاهش میدهد.
۳۲. تارهای عصبی به سه گروه اصلی A, B, C تقسیم میشوند. ویژگی اصلی هر گروه را بیان کنید.
نمایش پاسخ
پاسخ: گروه A: قطورترین تارها، دارای میلین ضخیم و بیشترین سرعت هدایت (مانند A-آلفا تا ۱۲۰ متر بر ثانیه).
گروه B: دارای میلین کمتر و نازکتر، سرعت متوسط (۳ تا ۱۵ متر بر ثانیه).
گروه C: نازکترین تارها، بدون میلین، کمترین سرعت هدایت (۰/۵ تا ۲ متر بر ثانیه) و آستانه تحریک بالاتر.
۳۳. «هدایت جهشی» (Saltatory Conduction) چیست و دو مزیت اصلی آن کدامند؟
نمایش پاسخ
پاسخ: در تارهای میلیندار، غلاف میلین عایق است و پتانسیل عمل فقط در گرههای رانویه (که فاقد میلین هستند) ایجاد میشود. به همین دلیل موج عصبی از یک گره به گره بعدی «میپرد».
مزایا: ۱. افزایش سرعت موج عصبی. ۲. صرفهجویی در مصرف انرژی (چون پمپ سدیم-پتاسیم فقط باید در گرهها فعال باشد).
۳۴. بیماری «ام اس» (MS) یا مالتیپل اسکلروزیس چیست و مکانیسم آن چگونه است؟
نمایش پاسخ
پاسخ: یک بیماری خود ایمنی است که در آن پادتنها و گلبولهای سفید خون به غلاف میلین حمله کرده و باعث التهاب و تحلیل رفتن آن میشوند. این امر موجب اختلال در هدایت امواج عصبی میگردد.
۳۵. «پتانسیل عمل مرکب» چیست و چرا از قانون «همه یا هیچ» پیروی نمیکند؟
نمایش پاسخ
پاسخ: پتانسیل عمل مرکب، پتانسیل عمل مجموع تارهای تحریک شده در یک عصب کامل (تنه عصبی) است.
این پتانسیل از قانون همه یا هیچ پیروی نمیکند، زیرا با افزایش شدت محرک، به تدریج تعداد بیشتری از تارهای عصبی (که آستانههای متفاوتی دارند) تحریک میشوند و در نتیجه ولتاژ کلی ثبت شده (پتانسیل مرکب) افزایش مییابد (پاسخ آن مدرج است).